Alternaria alternata is a necrotrophic pathogenic fungus, causing severe lost in many economic plants such as potato, pear, citrus, and tobacco. By using wild tobacco Nicotiana attenuata as a model plant, researchers in Kunming Institute of Botany find that host plants can defend themselves by closing their stomata by quickly activation of ABA signaling and mitogen activated protein kinase 4.
After infection, many fungal hyphae were observed to enter leaf tissue through stomata within 24 hours, and necrotic lesions were formed after 3 to 6 days. Importantly, source-sink transition leaves (0 leaves) were more resistant than all fully expanded ones, and this was correlated with more ABA and ABA-related gene transcripts and lower stomatal conductance in 0 leaves after infection. When supplied with ABA or incubated in the dark for one day, fully expanded leaves, which accumulated less ABA and showed higher stomatal conductance after infection in comparison to 0 leaves, showed increased their resistance. Mitogen-activated protein kinase 4-silenced plants, in which ABA-induced stomatal closure responses and resistance to A. alternata were highly impaired, did not show stomatal conductance change after infection, and both 0 and fully expanded leaves developed bigger lesions than in wild-type plants. Thus we conclude that ABA signaling pathway mediated by NaMPK4 is required for the resistance of N. attenuata to A. alternata, at least in part through the stomatal closure responses.
These results can partially answer the question why mature leaves are more susceptible to A. alternata in the tobacco field.
The story was published in the journal of “Plant Pathology”. All details were in http://onlinelibrary.wiley.com/journal/10.1111/%28ISSN%291365-3059/earlyview.
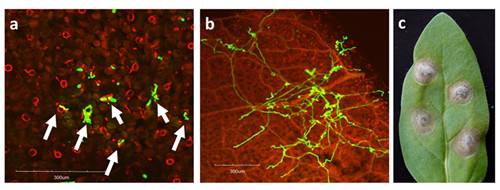
Figure1. Confocal microscopy observation of N. attenuata fully expanded leaf after A. alternata infection.
a: Penetration of the fungus starts at around 20 hpi;
b: Massive fungal growth can be observed at 48 hpi.
c: Disease symptom are fully developed at 5 dpi.
Leaves were inoculated as shown in Material and Methods. Fungal material was stained by WGA-AF488 (green). Plant material was stained by propidium iodide (red). White arrows indicate fungal penetration through stomata. Bars: 300 mm.
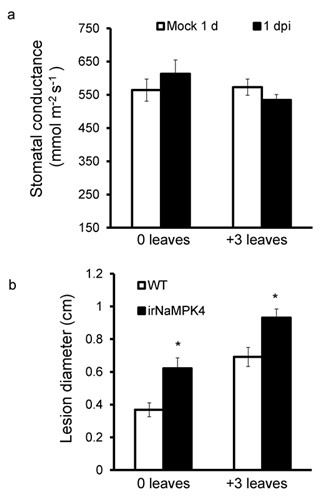
Figure 2 NaMPK4-silenced plants did not change their stomatal conductance after infection while both 0 and +3 leaves were more susceptible to A. alternata
a: Mean stomatal conductance measured in 6 biological replicated mock and A. alternata infected 0 and +3 leaves 1 dpi.
b: Mean diameter of necrotic lesions of 16 inoculation sites on 0 or +3 leaves from 4 different WT and irNaMPK4 plants. The experiment was repeated three times with similar results. The asterisks indicate significant differences between WT and irNaMPK4 plants (unpaired t-test: *, P<0.05).
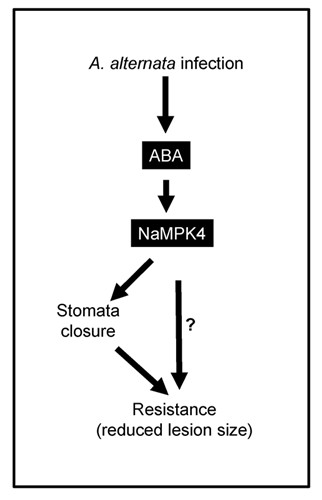
Figure 3. Working model of the role of ABA signaling pathway in N. attenuata-A. alternata interaction.
After A. alternata infection, ABA signaling pathway was activated in N. attenuata, and ABA exert its role through NaMPK4, and at least in part through stomatal closure response, finally affects the resistance of N. attenuata to A. alternata.



