Located in the centre of the Eurasian continent and representing the largest arid zone in the Northern Hemisphere, the arid Asian interior (here mainly referred to as Central and West Asia) is characterized by a continental climate with low annual precipitation and highly variable seasonal temperatures (Walter, 1979; Lioubimtseva and Cole, 2006).
Despite the harsh hydrothermal conditions, three regions of the Asia interior, i.e. the Caucasus, Irano-Anatolian region, and mountains of Central Asia, are listed among the 35 biodiversity hotspots defined by Marchese (2015).
During the past few decades, numerous studies have made major progress towards understanding the aridification processes and moisture patterns of the Asian interior (Miao et al., 2012; Hurka et al., 2019).
In contrast, the impact of aridification on the diversification process of plants inhabiting the interior Asia has been poorly documented from a phylogenetic and biogeographic perspective.
A close relationship between the floras of Central and West Asia and the Qinghai-Tibetan Plateau (QTP), another global biodiversity hotspot, has been suggested. Some lineages were inferred to have originated in the QTP and subsequently migrated to the inland Asia (the “out-of-QTP” hypothesis), whereas other lineages were purported to have originated in Central and West Asia and later dispersed to and diversified within the QTP.
This latter pattern is still poorly understood and requires additional documentation (Wen et al., 2014). The genus Dracocephalum L. (Lamiaceae, Nepetoideae, Mentheae) is an ideal group to investigate the evolution of the Asian interior arid zone flora and the relationship between the floras of Central and West Asia and the QTP, as the genus is most common in the steppe-desert biomes of the Asian interior and the alpine region of the QTP (Budantsev and Lobova, 1997).
During the past five years, the researchers collected materials of Dracocephalum and related genera from 15 countries in East Asia, Central Asia, Europa, and North America. Based on a sampling of 66 taxa comprising more than 80% from extant species of Dracocephalum s.l., morphological, phylogenetic (maximum parsimony, likelihood, and Bayesian inference based on nuclear ITS and ETS, plastid rpl32-trnL, trnL-trnF, ycf1, and ycf1-rps15, and two low-copy nuclear markers AT3G09060 and AT1G09680), molecular dating, diversification, and ancestral range estimation analyses were carried out.
As a result, two genera, Hyssopus and Lallemantia are embedded within Dracocephalum and nine well-supported clades can be recognized within Dracocephalum s.l. Analyses of divergence times suggest that the genus experienced an early rapid radiation during the middle to late Miocene with major lineages diversifying within a relatively narrow timescale. Ancestral area reconstruction analyses indicate that Dracocephalum s.l. originated in Central and West Asia and southern Siberia, and dispersed from Central and West Asia into the QTP and adjacent areas twice independently during the Pliocene.
The acidification of the Asian interior possibly promoted the rapid radiation of Dracocephalum within this region, and the uplift of the QTP appears to have triggered the dispersal and recent rapid diversification of the genus in the QTP and adjacent regions. Combining molecular phylogenetic and morphological evidence, a revised infrageneric classification of Dracocephalum s.l. is proposed, which recognizes nine sections within the genus.
The results were published online in the journal Cladistics entitled “Phylogeny and Biogeography of the northern temperate genus Dracoceaphalum s.l. (Lamiaceae)”, https://onlinelibrary.wiley.com/doi/10.1111/cla.12502
This work was supported by the National Natural Science Foundation of China (31800168), the Yunnan Fundamental Research Projects (2019FI009), the open research project of the Germplasm Bank of Wild Species, Kunming Institute of Botany, Chinese Academy of Sciences, the United States National Science Foundation (DEB-1655611), the Institute of Botany of the Academy of Sciences of the Republic of Uzbekistan (FZ-20200929321), and the Scientific Project of the State Order of the Government of Russian Federation to Lomonosov Moscow State University (121032500082-2).

Fig. 1. Morphological diversity and diagnostic characters of Dracocephalum s.l. (a) D. moldavica; (b) D. oblongifolium; (c) D. paulsenii; (d) D. peregrinum; (e) D. purdomii; (f) D. ruyschiana; (g) D. tanguticum; (h) Hyssopus officinalis; (i) Lallemantia royleanum; (j) calyces of D. argunense, showing the swollen folds at the base of the sinuses between the calyx teeth; (k) dissected corolla of D. bipinnatum, showing the two anther thecae diverging at 180° relative to each other; (l) bracts of D. rigidulum, showing the aristate teeth. a, c–h, j–l, photographed by Y.-P. Chen; b, photographed by T.O. Turdimatovich; i, photographed by N.Yu. Beshko. (Image by KIB)
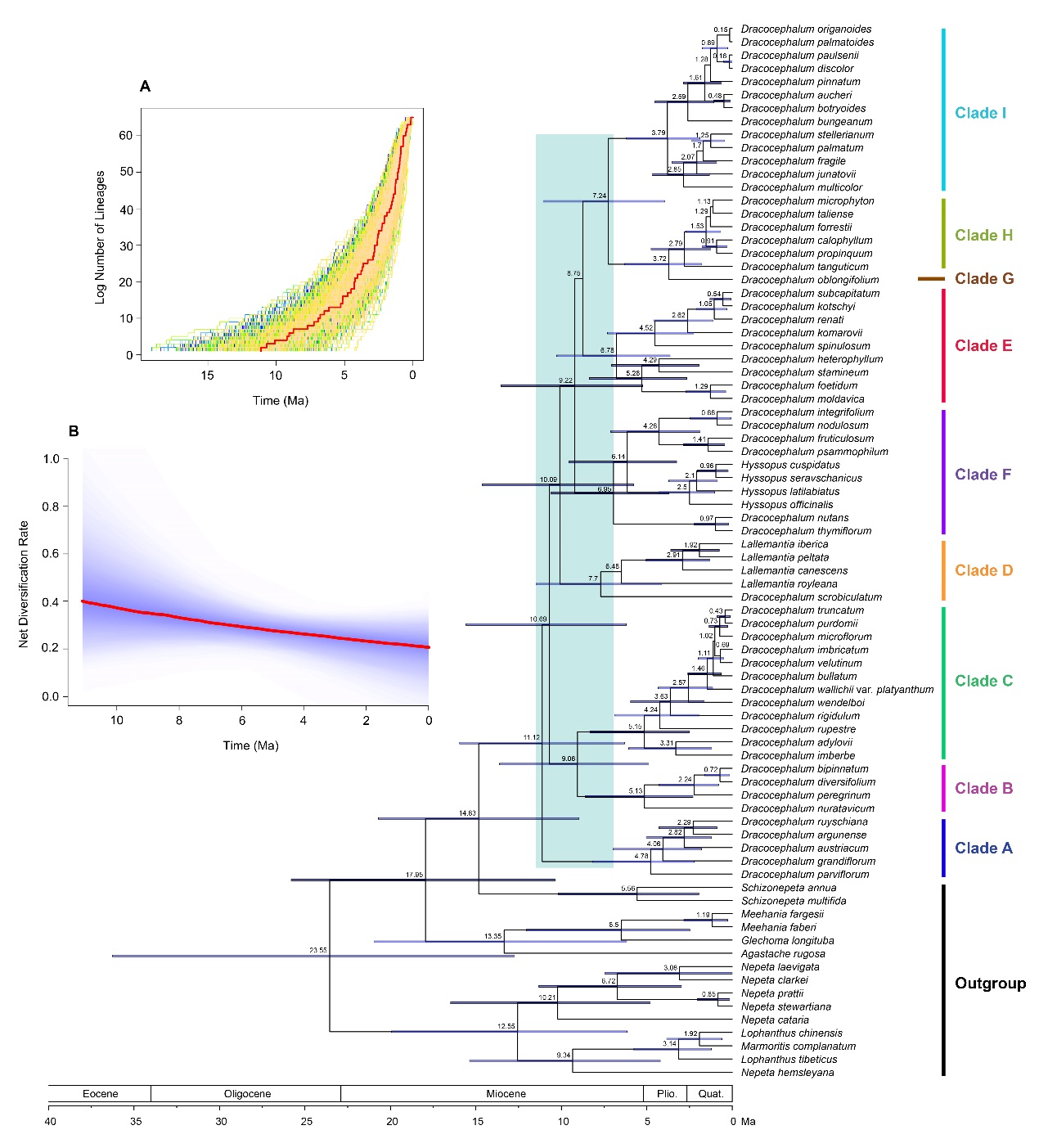
Fig. 2. Phylogenetic chronogram of Dracocephalum s.l. as inferred from BEAST analysis of the combined PPR data set. Blue bars represent 95% HPD of node age. An early rapid diversification of Dracocephalum s.l. occurred in late Miocene is indicated by green shading. Diversification dynamics of Dracocephalum s.l. are present at the upper left. (a) Lineage through time (LTT) analysis of Dracocephalum s.l. The red line indicates the MCC tree and the other lines represent 1000 random selected trees generated from BEAST analysis. (b) Rate-through-time of lineages according to BAMM. The red line represents the mean net diversification rate. Plio., Pliocene; Quat., Quaternary. (Image by KIB)
(Editor: YANG Mei)



